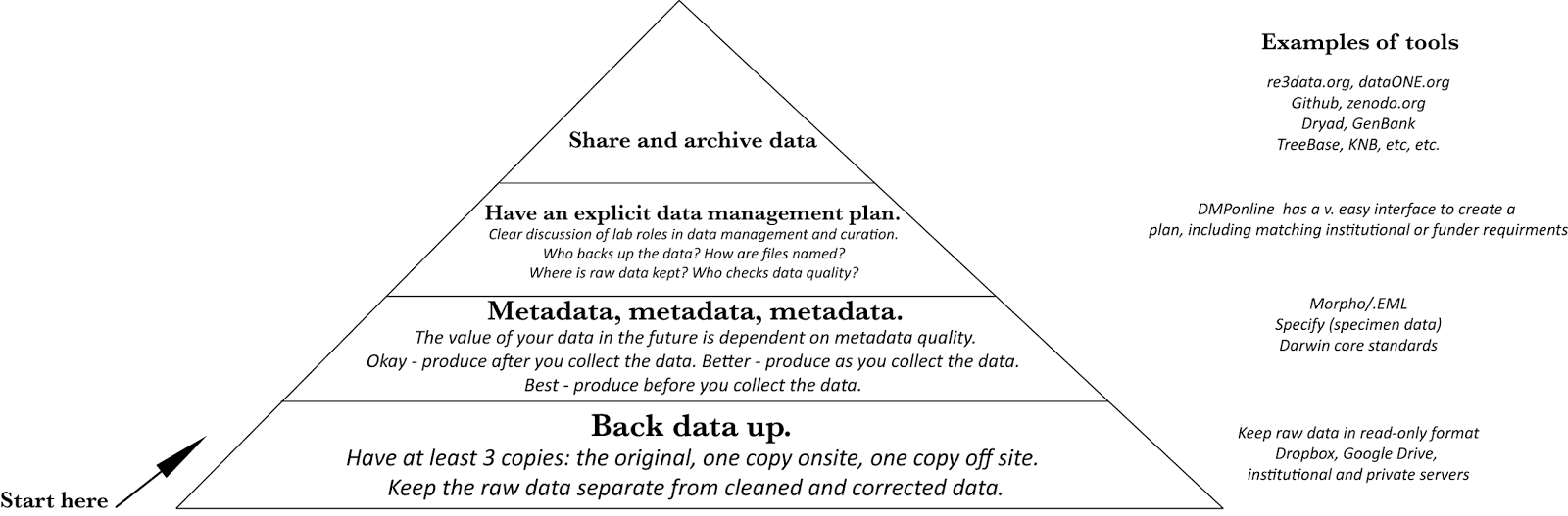
It seems at times that the focus on whether biodiversity has a positive relationship with ecosystem functioning has been a bit limiting. Questions about the BEF relationships are important, of course, since they support arguments for protecting biodiversity and suggests a cost of failing to do so. But as a hypothesis ('higher diversity is associated with higher functioning'), they can be rather one-dimensional. It's easy to think of situations in which other types of BEF relationships (neutral, negative) exist. So is it enough to ask if positive BEF relationships exist?
To test this, the authors estimated crown architecture for each species using traits that reflect crown shape and size. These measures were used to predict the spatial complementarity expected with different combinations of tree species. In addition, a single integrative trait – maximum growth rate – was measured for each species. The authors hypothesized that the variation in growth rate of species in a community would be associated with variation in crown heights and so also a good predictor of overyielding.
They found that crown complementarity occurred in nearly all of the experimental polycultures and on average was 29% greater in mixtures than monocultures. Controlling for the number of species, communities with greater variation in growth rate did in fact have greater crown complementarity, as predicted. Further, higher levels of crown complementarity were strongly associated (R2~0.6) with stem biomass overyielding.
These results provide a clear potential mechanism for a positive effect of biodiversity (particularly trait-based variation) in similar forests. (As they state, "We posit that crown complementarity is an important mechanism that may contribute to diversity-enhanced productivity in forests"). Given the importance of the sun as a limiting resource in forests, the finding that mixing species that combining shade intolerant and shade tolerant strategies are more productive (the authors note that "growth rate aligns with shade tolerance and traits indicative of a tree’s resource strategy") is not necessarily surprising. It fits within existing forestry models and practices for mixed stands. This is a reminder that we already understand many of the basic components of positive (and neutral and negative) diversity-functioning relationships. The good news is that ecology has accumulated a large body of literature on the components of overyielding (limiting resources, niche partitioning, evolution of alternate adaptive strategies, constraints on these, the strength of competition, etc). From the literature, we can identify the strongest mechanisms of niche partitioning and identify the contexts in which these are likely to be relevant. For example, sun in forests and canopy complementarity, or water limitation in grasslands and so root complementarity might be a good focal trait.
It’s nice then that there is increasingly a focus on identifying mechanisms behind BEF relationships, using both theory and empirical research. A new paper along these lines is “Spatial complementarity in tree crowns explains overyielding in species mixtures” from Laura Williams et al. (2017). "Overyielding" is the phenomenon in which greater total biomass is produced in a mixture of species compared to the expectation based on their biomass production in monoculture. Overyielding would suggest a benefit in maintaining polycultures, rather than having monocultures, and is a common response variable in BEF studies.
This study focused on the production of stem biomass in monocultures vs. polycultures of forest trees. Experimental communities of young tree species were planted with orthogonal gradients of species richness and functional richness, allowing the effects of species number and trait diversity to be disentangled. Complementarity in tree canopy structure in these communities may be an important predictor of overyielding in stem biomass. Complementarity among tree crowns (that is, the extent to which they fit together spatially without overlapping, see Fig below) should reflect the ability of a set of species to maximize the efficiency of light usage as it hits the canopy. Such variation in crown canopy shapes among species could lead to a positive effect of having multiple species present in a community.
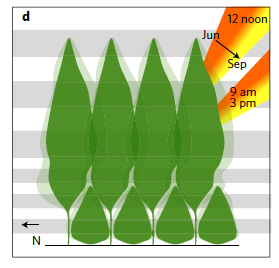 |
| Example of crown complementarity. From Williams et al. 2017. |
To test this, the authors estimated crown architecture for each species using traits that reflect crown shape and size. These measures were used to predict the spatial complementarity expected with different combinations of tree species. In addition, a single integrative trait – maximum growth rate – was measured for each species. The authors hypothesized that the variation in growth rate of species in a community would be associated with variation in crown heights and so also a good predictor of overyielding.
They found that crown complementarity occurred in nearly all of the experimental polycultures and on average was 29% greater in mixtures than monocultures. Controlling for the number of species, communities with greater variation in growth rate did in fact have greater crown complementarity, as predicted. Further, higher levels of crown complementarity were strongly associated (R2~0.6) with stem biomass overyielding.
 |
| Fig 2&3 from Williams et al (2017). For experimental communities: a) the relationship between crown complementarity and variation in growth rate. b) the relationship between crown complementarity and stem biomass overyielding. |
These results provide a clear potential mechanism for a positive effect of biodiversity (particularly trait-based variation) in similar forests. (As they state, "We posit that crown complementarity is an important mechanism that may contribute to diversity-enhanced productivity in forests"). Given the importance of the sun as a limiting resource in forests, the finding that mixing species that combining shade intolerant and shade tolerant strategies are more productive (the authors note that "growth rate aligns with shade tolerance and traits indicative of a tree’s resource strategy") is not necessarily surprising. It fits within existing forestry models and practices for mixed stands. This is a reminder that we already understand many of the basic components of positive (and neutral and negative) diversity-functioning relationships. The good news is that ecology has accumulated a large body of literature on the components of overyielding (limiting resources, niche partitioning, evolution of alternate adaptive strategies, constraints on these, the strength of competition, etc). From the literature, we can identify the strongest mechanisms of niche partitioning and identify the contexts in which these are likely to be relevant. For example, sun in forests and canopy complementarity, or water limitation in grasslands and so root complementarity might be a good focal trait.